I. En arvelig mekanisme som bevarer celle-type
Hvordan kan utrolig presise mønstre for hvordan gener uttrykker seg, overføres innen ulike celletyper, når DNA-sekvensen forblir den samme i alle? Det kan sammenlignes med at hver celletype får sine egne typer script, som er tilpasset deres egen rolle. En egen mekanisme sikrer konsistens, i hvordan genuttrykk overføres, og gjør at hudceller fortsetter å produsere hudceller, ikke andre celletyper.
Spørsmål å reflektere over:
Hvordan kan en slik mekanisme utvikles innen cellers livsløp?
Hvordan kan genetisk like individer framtre så forskjellige?
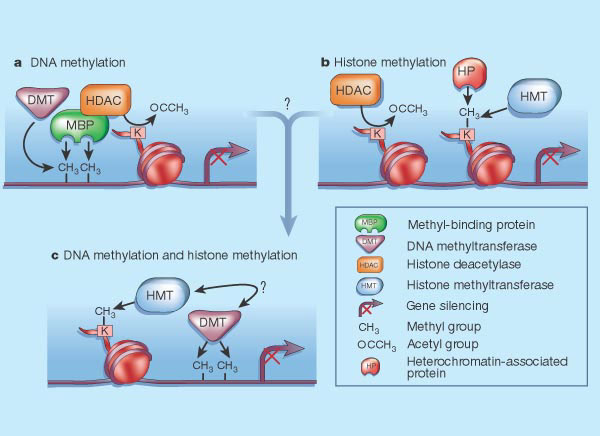
Bilde 1A. Metylert DNA og histoner
Bilde 1B. Genetisk like mus- med ulikt utseende |
  |
II.A Det foreligger samsvar og samarbeid mellom ulike nivåer genetisk og epigenetisk
Metyltaggers binding til cytosin må være fininnstilt og samstemt. Enzymer danner epigenetisk kode, som gir ny kjemisk struktur, og endrer virkemåte. Denne er nødvendig for at organismer skal fungere. For å skjønne hvordan dette foregår, trengtes de skarpeste hjerner innen biologien. . Algoritmer er kodet og lagret i DNA. Spørsmålet er hvordan alle ledd i denne rekken kunne danne funksjonelle koder for levende organismer. 'Mind' virker etter intelligente mønstre, mens 'matter' vil virke enten tilfeldig eller lovbestemt. |
 |
II.B. Kompleks og spesifisert kausalitet i over 200 ulike menneskelige celletyper
Det store flertall av forskere holder nå på at epigenetiske endringer forårsaker endringer i hvordan gener uttrykkes (kausalitet). Tidligere mente noen at gener kunne undertrykkes av en ukjent mekanisme, som så gjorde at DNA ble metylert, og skrudde av geners virkemåte (mer som form av 'uhell'). Mønstre av DNA-metylering er ulikt over ulike celletyper.. Bilde 2. Acetyl-tagging på histon-spoler |
III. Enzymers metylering danner kaskade-reaksjoner, som avhenger av hvor omfattende metyleringen er.
Proteiner som bare bindes til metylert cytosin, setter cellen i stand til å fortolke endringer i DNA-regionen, og fungerer som lesere av epigenetisk kode.
Det er en merkelig og udokumentert påstand at en slår fast eksistensen av en sammenhengende rekke av naturlige, ikke-styrte funksjonelle, genetiske kaskade-reaksjoner, fra urcelle til menneske.
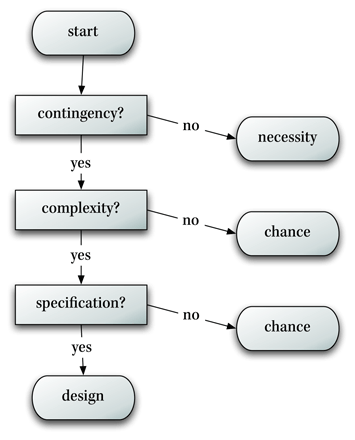
William Dembski greide ut om design-slutninger i hva han kalte et forklarings-filter: "Lovmessigheter (contingency) er alltid 1.linje-forsvar. Om vi kan forklare ved hjelp av det, er tilfeldighet og design utelukket. På lignende måte er tilfeldighet 2.linje-forsvar. Om vi ikke kan forklare et fenomen ved lovmessighet, men ved tilfeldighet -er design utelukket. Det er en prioriteringsrekkefølge, der lovmessighet kommer først, så tilfeldighet og dernest design.
Det vanskeligste å ta stilling til, er hvorvidt tilfeldighet er involvert: Dersom noe ikke er komplekst nok til å kunne være design, skyldes det tilfeldigheter. Det kan det også gjøre, selv om det er komplekst -bare det ikke er spesifisert. Det tredje steget i filteret er der for å hindre design-slutning i slike tilfeller. Det gjøres ved å benytte et begrep som kalles spesifikasjon (specificiation). En spesifikasjon er et gjenkjennbart mønster, som eksisterer uavhengig av fenomenet vi analyserer. Dembski forklarer: det er bare når den presise sekvensen av forekomster er blitt tilsagt på forhånd, at vi har grunnlag for å tvile om dette skjedde tilfeldig. En kan uttrykke det som at begivenheten stemmer med et mønster, som leder oss til å lete utenom tilfeldigheter for å forklare begivenheten.
Bilde 3. Dembskis-filter
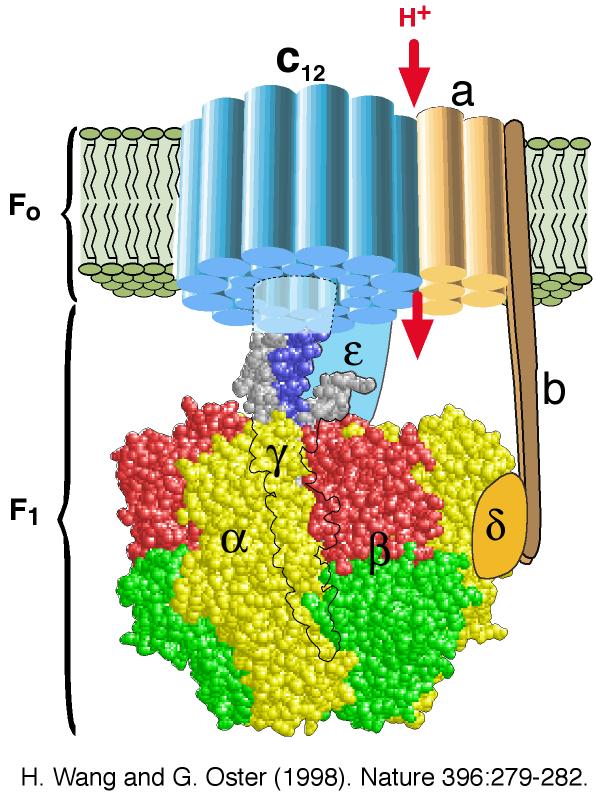
Bilde 4. ATP-motoren, eks. på spesifisert kompleksitet |
  |
IV. 'Enigma-koden' har vært avdekket og forstått -lenge
DNA-metylering er helt nødvendig, og må ha vært tilstede fra starten av høyere organismers eksistens. Hjerneceller produserer verken hemoglobin eller fordøyelses-enzymer f.eks. For å hindre prosesser som er vanlige i visse andre celler, men som ville vært dødelige i hjernen, har DNA-metylering holdt visse deler av genomet fullstendig tillukket.
For hudceller derimot, så må disse stadig kopieres. Om de er metylerte, finnes (DNMT1-) enzymer, som kan gjenkjenne hvorvidt Cytocin er metylert, og tilfører en tilsvarende metylering på nylig kopiert DNA-element.
Genekvenser fra sjimpanse kan inneholde bearbeidede, repeterende enheter, som kalles palindromer, der innholdet er det samme -om det leses forfra eller bakfra!
Bilde 5. DNA-metylering |
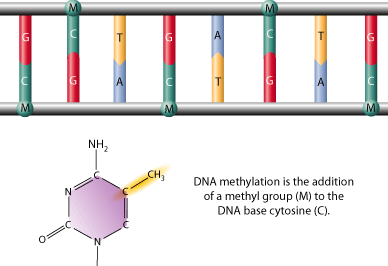 |
V. Det overordnede nivåets betydning er avgjørende for liv og helse- mens mutasjoner bryter ned
Det er gjort epi-genetiske forsøk med mus der kurering av RETTs syndrom har funnet sted. Det forårsakes av en mutasjon i MeCP2-genet. Det er genet som koder for lesning av metylert DNA. Barn med dette syndromet produserer ikke et fungerende MeCP2 protein, fordi cellene deres ikke kan lese denne delen av epigenetisk-kode skikkelig. Det ble innsatt en normal, men stilnet kopi av fungerende Mecp2, i en mus-unge som hadde arvet det muterte MeCP2-genet.
Da man (Adrian Bird) klarte å slå på det fungerende Mecp2, ga det helt tydelige og synlige resultater i form av økt nysgjerrighet og aktivitet fra mus-ungen.
Bilde 6. Mus kurert fra RETTs-syndrom |
 |
VI. Nukleotider og nukleosomer i mikro-kosmos
En menneskecelle er ca. 1/100 millimeter i diameter. DNAet til verdens befolkning vil kunne få plass i en teskje.
Bilde 7A. Nukleotide -til venstre:
DNA er intimt assosiert med proteiner, og spesielt med proteiner som kalles histoner. Det er fire ulike typer (H2(A og B), H3 og H4). De har en 3 dimensjonal globus-form, og er formet i kompakte ball-lignende former. To kopier av hver av disse kommer sammen i en åtte-gruppert sammensetning.
Se Bilde 7B.Nukleosom -til høyre
147 basepar av DNA er kveilet rundt hver nukleosome. DNA-regionene er tungt metylerte rundt disse hyper-kondenserte strukturene, noe som benyttes for å holde visse gener avslått gjennom tiår i lenge-levende nevroner. Nå finnes det gener som har potensiale til å bli slått på. Histon-modifiseringer er mer foreløbige modifiseringer enn DNA-metylering. De fungerer som tilpassede script, som kan gjennomleve noen omganger med kopiering, eller er meget midlertidige. |
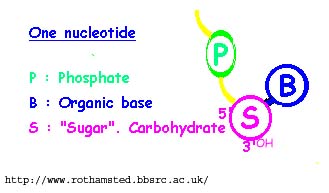  |
VII: Fininnstilling av liv
I det utviklende embryoet binder proteiner seg til gener og slår disse på. Noen av disse genene kan selv kode for epigenetiske enzymer, som vil danne komplekser av medlemmer av en ny genfamilie, osv. De genetiske og epigenetiske proteinene arbeider sammen i en 'sømløst' ordnet progresjon. Noen ganger vil en celle uttrykke litt mer eller mindre av en nøkkel-faktor, hvis verdi er fininnstilt. Det kan i sin tur endre utviklingsstien som cellen tar. Små endringer i hvordan 'bitene ble tilpasset' vil trigge én utvikling, og ikke en annen.
Geninnstillinger er mye mer finjusterte enn å være AV eller PÅ. De er mer like en volum-knapp på en radio. Det finnes faktisk mer enn 50 ulike epigenetiske histon-modifikasjoner (1). Mønsteret i modifisering refereres til som histon-kode.(2) Det er en kode som har vist seg meget vanskelig for epigenetikere å lese. Men selv om man ikke p.t. har klart å tyde den, jobbes det intenst med saken i laboratorier verden over, fordi man skjønner at den er ekstremt viktig.
Fininnstilling av Carbon-produksjon: Bilde 8.A. 3-alfa-prosess og
Bilde 8B. Ikke-karbon basert liv er trolig science-fiction |
  |
VIII. Et 'timet og tilrettelagt' sam-utviklingsprogram
Det utviklende embryoet er å likne med et kaskade-spill. Zygoten er ferdiglastet med visse proteiner, mest fra eggets cytoplasma. Disse proteinene går inn i kjernen av zygoten og bindes til mål-gener. De tiltrekker også noen få epigenetiske-enzymer til disse genene. De epigenetiske enzymene setter opp mer langvarige endringer til DNA og histon-proteiner, som influerer hvordan genene slås av og på.
I tidlig embryo, forekommer feed-back. Ett av nøkkel-proteinene i det tidligste embryonale stadium, er Oct4. Det binder seg til spesielle nøkkel-gener. Både Oct4 og det epigenetiske enzymet det jobber sammen med er helt nødvendige for utvikling. Uten at begge er til stede kan ikke zygoten en gang starte utviklingen. Oct 4 er benyttet som en reprogrammerings-faktor, der celler ble narret til å opptre som tidlige utviklings-celler. Selv epi-genetiske modifikaterer ble resatt, så viktig er Oct4-genet. Menneskelig sykdom kan også framkalles av mutasjoner i enzymer som fjerner epigenetiske mutasjoner(PHF) -(3).
I løpet av svangerskapet dannes i gjennomsnitt millioner av celler pr. sekund. Et biologisk ur styrer utvikling av nye kroppsceller. Senere i livet nydannes hvert sekund ca. to millioner røde blodceller (lenke), for å holde organismen ved like.
Bilde 9. Gastrulasjon av zygote |
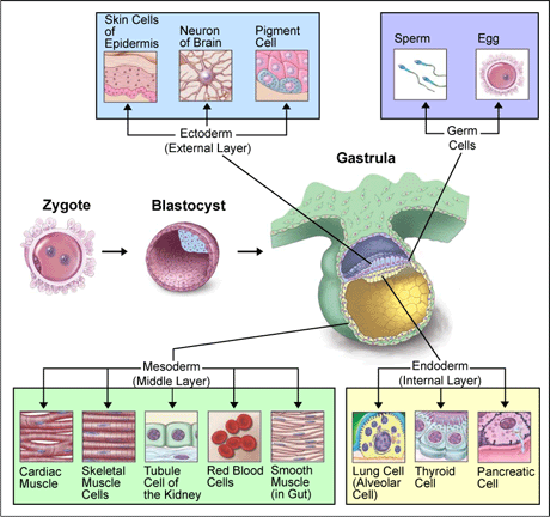 |
IX. Cellens innfløkte informasjonssystem
I tillegg til disse histon-'lesere' og 'slettere', så eksisterer det over 100 proteiner, som fungerer som lesere' av denne histon-koden, ved å binde seg til epigenetiske-markører. Disse 'leserne' tiltrekker proteiner og bygger opp komplekser, som skrur av eller på genuttrykk. De fleste histon-modifikasjoner er mye mer responsive enn DNA-metyleringer. Metylert DNA har en tendens til å forbli slik under de fleste forhold. Det er derfor nevroner forblir nevroner, og det er temmelig uvanlig at DNA-metylering fjernes i celler.
Histon-kode kan derimot endres som respons på hormoner, f.eks. insulin som signaliserer til muskel-celler, eller østrogen som påvirker bryst under menstruasjonsperioder. Næring i omgivelsene kan samvirke med våre gener via endringer i histon-koden, men grunnlaget for slike endringer må være innebygd i cellen.
|
 |
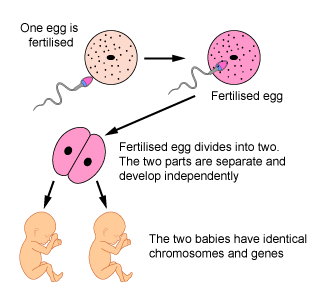
X. Eneggede tvillinger, ikke identiske: Mer enn gener
I 2005 ble det offentliggjort en artikkel av en stor gruppe ved det Spanske nasjonale kreft-senteret, ledet av Professor Manel Esteller. Da de undersøkte én-eggede tvillingpar, kunne de ikke se mye forskjell i nivåer av DNA-metylering eller histon-acetylering mellom de to tvillingene. Men da de undersøkte par av én-eggede tvillinger som var mye eldre, var det mye variasjon mellom tvillingene i så måte. Spesielt syntes dette å være tilfelle hos tvillinger som hadde levd fra hverandre i lang tid.
Ett team ved Royal Chiildrens hospital i Melbourne undersøkte DNA-metylering hos én- og to-eggede tvillinger. De undersøkte noen få områder av genomet i større detalj enn Esteller (4). I tillegg til relativt store ulikheter i DNA-metylerte mønstre hos to-eggede tvilling baby-par, fant de at også én-eggede tvillinger avvek fra hverandre, i DNA-metylerte mønstre. Dette indikerer at én-eggede tvillinger begynner å divergere epigenetisk, mens de befinner seg i livmoren.
Vi kan konkludere at selv genetisk identiske individer er epigenetisk distinkte i fødselsøyeblikket, og disse epigenetiske forskjellene blir mer uttrykt etter hvert, særlig om de blir utsatt for ulike omgivelser.
Bilde 11A. Eneggede tvillinger Bilde 11B. -kan være epigenetisk ulike ved fødsel
|
  |
XI. Om mus og menn -ulik inngang for forskere
Hos mus har en funnet en egenskap hvor halen er vridd, som viser stor variasjon, selv mellom genetisk identiske individer. Det har vist seg å være ett eksempel der variasjonen skyldes ulike nivåer av DNA-metylering, innen ett hoppende gen i ulike dyr.
Selv om det ikke er aktuelt å forske direkte på mennesker, så har likevel omstendighetene en sjelden gang gjort at det er påvist klare sammenhenger mellom at uheldige miljømessige påvirkninger for fostre i tidlig embryo-periode, varig påvirker livskvalitet.
Ett uår som virker inn på svangerskapets tidligste fase, kan få komplikasjoner som varer over tiår. Dette er konsistent med at det er da mønstre setter seg, i form av at genuttrykk preges via epigenetiske metyleringer. Nylige studier har vist endringer i nøkkel-gener, involvert i stoffskifte. Dataene er konsistente med at underernæring i tidlig utviklingsperiode, endrer epigenetisk profil hos nøkkel-gener knyttet til stoffskifte (5). Cellene synes å tilpasse seg som best den kan, under forhold med dårlig næringstilgang, men negative virkninger synes å akkumulere seg på sikt.
Bilde 12A. Ulik DNA-metylering hos genetisk like mus
|
  |
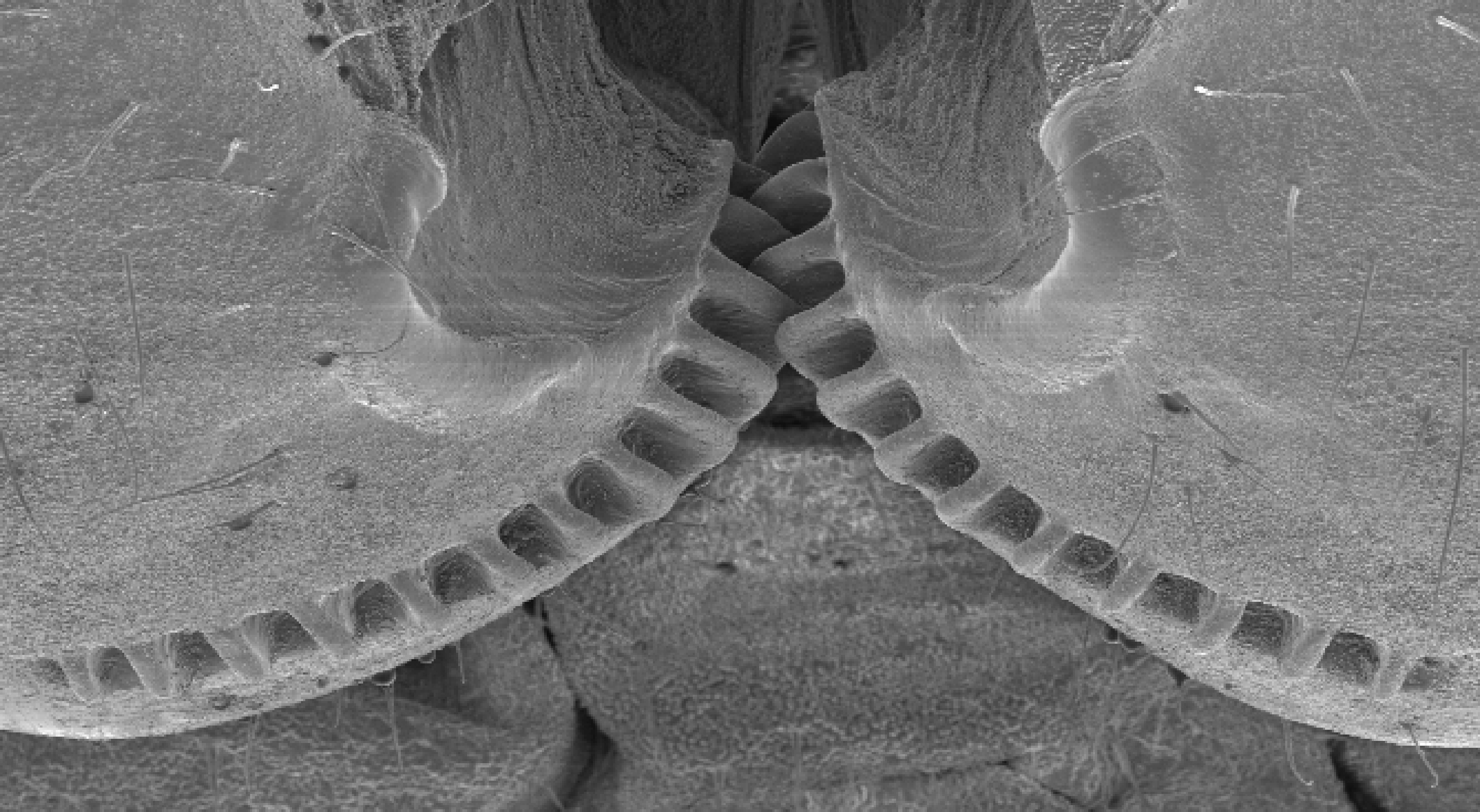
XII. En ung skjoldsikade (Issus coleoptratus) kan akselerere fra 0 til ca. 4 m/sek på to tusendedeler av et sekund. Det utsetter kroppen for et trykk som er nesten 400 ganger så kraftig som gravitasjonskraften. Slike hopp krever at insektet sparker like hard fra med begge ben nøyaktig samtidig.
Grunnen til at skjoldsikaden kan gjøre dette, er at innerst på hvert av de to bakbeina, har de en rad med avrundede tannhjultenner som griper inn i hverandre og kobler bena sammen. Uten disse ville ikke hoppet vært synkronisert og insektet ville skjene av gårde uten styring.
I det siste hamskiftet før voksenstadiet forsvinner disse tannhjulene. Innen da synes de å ha lært å 'styre seg'. Bilde 13.A. Tannhjul hos skjoldsikade (Issus -kredit: Burrows/Sutton)
|
  |
{kind=link}
{kind=link}
{kind=link}